

Over the course of evolution, cells have internalized factors in their environment that have posed existential threats..
Introduction
How and why evolution works has proven refractory to experimentation- how fish have evolved into Man, for example- because evolution is thought to be due to random mutations, obviating the opportunity to test such a mechanism, leaving only correlations and associations, which do not show causation. Ideally, there would be some comparable process that could be exploited in order to deconvolute evolution, but none has been forthcoming up until now, having been introduced to Knot Theory, a subheading of Topology. Topology concerns itself with how geometric objects maintain themselves under continuous deformations. Knot Theory is the study of mathematical knots, inspired by the knots that occur in everyday life. It occurred to us that since knots tie things together, they are similar in kind to the cell-cell signaling mechanisms that tie physiologic traits together during development, culminating in physiology, ultimately tied together by homeostatic forces.
Evolution is topologic because it began as a ‘surface’, formed by lipid micelles submersed in the ocean that covered the primordial earth, forming a boundary between the Cosmos and the interior of micelles. Such protocells comply with the First Principles of Physiology- negative entropy, chemiosmosis and homeostasis. Negative entropy runs counter to the positive entropy in the environment, forming an ambiguous state of being. The processes of development and phylogeny inform the organism of environmental changes, the unicell twisting and turning in its efforts to conform with its environment in order to act as a phenotypic agent, collecting epigenetic ‘marks’ that keep the organism informed of environmental changes. Such data are then assimilated by the egg and sperm in order to inform the offspring as to how and why to adapt.
In the case of mathematical knots, they are also generated from cell-like circles that ‘twist and turn’, held together by theoretical ‘springs’, and can be untied by releasing them, re-forming as circles. In the case of physiologic knots, they ‘untie’ themselves in response to physiologic stresses, ultimately during the process of aging or senescence, returning to the unicellular state as a ‘circle’ over the course of the life cycle.
The Homology Between Evolution and Knots
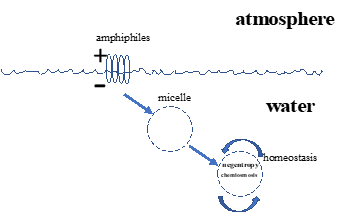
| Fig. 1. Formation of Micelles from lipids immersed in water. Lipids immersed in water initially align perpendicularly to the surface as amphiphiles, their hydrophobic ends facing upward, their hydrophilic ends facing downward. These lipids will then spontaneously form micelles, spheres with semi-permeable membranes surrounding them because the lipids reduce the otherwise high surface tension of water. tension. |
As stated in the Introduction, knots are surfaces, which is how they relate to evolution- lipids originating either from Pulsars or from thermal vents in the sea floor localizing to the air-water interface [Fig. 1], where they formed micelles, prototypical cells that formed a surface between the Explicate and Implicate Orders, forming what Claude Bernard referred to as the milieu interieur. This is the origin of unicellular organisms, which dominated the earth for the first 3.5 billion years. Due to competition between prokaryotes and eukaryotes, the latter began forming multicellular organisms through cell-cell communications mediated by growth factor-growth factor receptor signaling mechanisms. Such cellular pathways for growth and differentiation during embryologic development subsequently become the homeostatic basis for physiology, as alluded to above. In the process of doing so the cells involved produce extracellular matrices that stabilize them physically, forming true ‘knots’ that literally tie our physiology together as organisms. During processes of chronic disease, such cellular communications break down, reverting back to earlier developmental and phylogenetic homeostatic stages of evolution, stabilized by scar tissue. During the aging process, loss of bioenergy similarly leads to systematic loss of homeostatic control and deterioration in cell-cell signaling, revealing the knot-like characteristics of organismal structure and function.

| Fig.2. Endosymbiogenesis. Over the course of evolution, cells have internalized factors in their environment that have posed existential threats, represented by x, y and z. The compartmentalization of such factors is the basis for cellular physiology. |
Over the course of vertebrate evolution, organisms have coped with their ever-changing environments by endogenizing factors in their environment- heavy metals, gases, ions, bacteria- referred to as the Symbiogenesis Theory [Fig. 2]. Once such factors were internalized, they were compartmentalized using intracellular membranes, forming what is commonly referred to as cellular physiology. Organisms became progressively more and more complicated based on cellular communications, producing the phylogenetic changes of speciation. But it should be borne in mind that organisms always return to the unicellular state over the course of their life cycle. The capacity to form and unform structure and function, all the way back to a ‘circle’ is also the property of a mathematical knot.
Circles, Knots, Cells and Memory
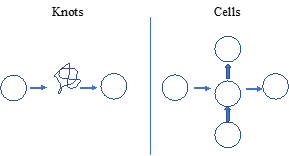
| Fig. 3. Relationship between topologic ‘knots’ and cellular evolution. On the left is a schematic for knots as simple circles that can then for complex configurations, but can be un-knotted to reform as circles. On the right is a depiction of cellular evolution, beginning with the unicell (left), forming complicated organisms through vertical integration of cell-cell signaling (middle), ultimately reverting back to the unicellular state (right). |
Memory is essential to the process of evolution- in order to remain in sync with an ever-changing environment the organism must remember its past ways of adapting as the most efficient way of surviving, referred to as exaptations. It is commonly held that DNA is our biologic form of memory, but the use of nucleic acids for this purpose is derivative of the lipid-based micelle, which has molecular memory in the form of hysteresis. The capacity to deform when warmed by the Sun, and re-form in the cold of night is the origin of memory. Like this most basic functional property of life, knots also have memory, because as complicated as they may become as Trefoils, Borromeans, etc, they remain simple circles at their origin as the formal validation that they are in fact knots [Fig. 3]. The same holds true for a complicated organism, because it ultimately returns to its unicellular form over the course of its life cycle, proving that it, too, is a knot. By returning to the unicellular state, the organism provides a way of integrating epigenetic ‘marks’ into its physiology over the course of ontogeny, explaining the cellular basis for evolution.
And short of regressing to the unicellular state, tissues can default to earlier stages in their ontogeny and phylogeny due to loss of cell-cell signaling. This mechanism is due to the cellular networking for development and homeostasis, and it provides the option of reverting to an earlier physiologic state in order to maintain equipoise, allowing the organism to reproduce and transfer its genome to the next generation.
DNA Knots, Homeobox Genes and Cell-Cell Signaling
Evidence that DNA ‘knots’ are altered by changes in cell-cell signaling is not forthcoming. Yet we know that under various stress conditions, cells produce Radical Oxygen Species that may cause mutations and duplications. Such DNA changes are then acquired by the genome, hypothetically altering the topology of DNA. The best evidence for this lies in the fact that Homeobox gene DNA is arrayed along chromosomes in the same way that it appears in the organism.
Pre-adaptations, Lipids Immersed in Water, and Knots
The cellular approach to evolution is predicated on serial pre-adaptations or exaptations, raising the question as to where and when the knots for life began? In the scenario for the role of lipids in water as the genesis of life, lipids on earth arose from both thermal vents in the sea floor as mentioned earlier, or as hitch-hikers on the asteroids that delivered frozen water to earth, the lipids originating from Pulsars.
Initially, the lipids aligned perpendicularly to the water-atmosphere interface [Fig.1] because they are polarized amphiphiles, their positively-charged ends facing upwards, being hydrophobic, their negatively-charged hydrophilic ends immersed in the water phase. In that configuration they behave like antennas, responding to electromagnetic forces such as Pulsars and photons. When the lipids form micelles, which are spheres with semi-permeable membranes, the effect of the electromagnetic force acts to stimulate cytoplasmic streaming as a phenotypic flow of energy. Subsequently, when cholesterol appeared in the cell membrane, it caused thinning of the membrane, further facilitating cytoplasmic streaming as locomotion. Fast-forward to Ciona intestinalis, a unicellular organism in which the stem cells for the heart develop in the tail, and then migrate into the body to form the heart. It is within the realm of possibility to think that the beating of the tail translates into the beating of the heart. This process may have exaptively evolved from the earlier evolution of cytoplasmic streaming since the heart evolved as a muscularization
of the vasculature, like the heart of an earthworm, perhaps due to the homologous ‘cytoplasmic streaming’ of blood acting to cause shear stress, known to generate gene mutations and duplications by generating Radical Oxygen Species [Fig. 4]. The one-chambered heart of the earthworm could have given rise to the two-chambered hearts of fish, the three-chambered hearts of amphibians and reptiles, and ultimately to the four-chambered hearts of birds and mammals due to beta-adrenergic agents like
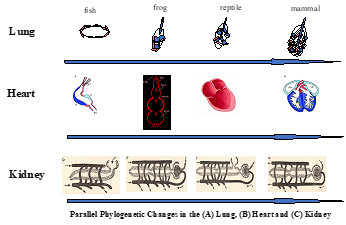
| Fig. 4. Evolution of the lung, heart and kidney. The evolution of the lung (top), heart (middle) and kidney (bottom) is depicted based on developmental and phylogenetic data. Note the increasing complexity of these phenotypic traits from left to right due to the interaction of a changing environment and the cellular components of each structure/function based on cell-cell signaling controlled by homeostasis. |
adrenaline. If you delete the beta-adrenergic receptor from developing mice, for example, they are born with a two-chambered heart. The beta-adrenergic receptor ‘duplicated’, i.e. amplified during the water-land transition, likely promoting the evolution of the heart from two chambers in fish, to three chambers in amphibians and reptiles, to four chambers in birds and mammals.
The above provides a ‘knot’ that begins with lipids forming a boundary between the Explicate and Implicate Orders, all the way through to the evolution of the lung, heart and kidney. The genetics of heart development, for example, are connected to those of the fingers, exemplifying the phenotypic homology between these structures based on cytoplasmic streaming.
The Deep Significance of the Homology between Knots and Evolution
As expressed above, the mechanism of cellular evolution is predicated on exaptations from sequentially earlier and earlier genetic traits, working backwards from present-day physiology to the unicellular state. Starting with lung evolution proved to be fortuitous because gas exchange is a very ancient property of life that allowed tracing of the process all the way back to the unicell. But that begs the question as to what the prototype for the cell itself was? The only such unity seems to be the putative Singularity of nature that existed prior to the Big Bang. The rationale is that the unicell has the capacity to express all of the states of being- homeostasis, mitosis and meiosis- all three being determined by the cytoskeleton as the ‘sensor’ for the prevailing environmental conditions. But the insight that knots and evolution share common properties suggests that perhaps even before the homology between the cell and the Singularity, the shared topology of knots and biology may have formed the ultimate homology of the inanimate and animate.
The Metaphysics of Knots
The homology of physical and cellular knots provides insight to the continuum from the inanimate to the animate. The phenomenon of lateralization extends that even deeper into our psyche by realizing that our right and left cerebral hemispheres act to perceive the left and right sides of our bodies, respectively, perhaps ‘unknotting’ the ambiguity from which we originated
due to the negative entropy that Schrodinger described in What is Life. By crossing over from left to right, or right to left in perceiving our surroundings we are able to reconcile that ambiguity and the subjectivity of our senses it gave rise to evolutionarily, seeing the
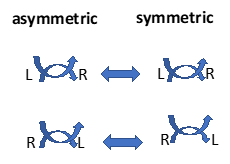
| Fig.5. Lateralization. We perceive things through the asymmetry of our left and right cerebral hemispheres (on the left). However, when we look one another in the eyes, those asymmetries are reconciled (on the right). |
environment in what David Bohm referred to as the Implicate Order [Fig.5].
However, when we look one another in the eyes, we transcend that ‘trompe l’oeil’. We instinctively recognize this when a mother looks into her newborn’s eyes, as love at first sight, or when we dance together. Perhaps this is what happened to Narcissus when he saw his reflection in a pool of water? Or when an artist conveys a novel or a painting or a piece of music? I have experienced insights from time to time while shaving…..perhaps it was due to looking in the mirror while holding cold steel to my throat that gave clarity, like what I am expressing here?
Discussion
Gene mutations as the cause of Darwinian evolution dissociate that process from development and phylogeny as sequential processes. Conversely, the reverse-engineering of cell-cell communications has offered fundamental insights to the ‘how and why’ of the ontology and epistemology of evolution. The current delineation of the interrelationship between cell-cell communication and Knot Theory further expands on such fundamental causal mechanisms, providing support for the previously expressed concept of “The Singularity of Nature” (Torday, 2019). It is only through such cross-cutting, transcendent diachronies that we will ultimately be able to form an algorithmic perspective for a holistic understanding of life in the Cosmos.
Up until now it has only been the artists who have been able to express the holism of our existence. Frost said that ‘life is the which can mix oil and water’, and that ‘The Secret Sits’ in the middle, and we dance around wondering why. Similarly, in Little Gidding, T.S. Eliot assures us that
“We shall not cease from exploration
And the end of all our exploring
Will be to arrive where we started
And know the place for the first time.”

| Fig.6. Magritte’s “Not to be reproduced” |
| René Magritte. La Reproduction interdite (Not to Be Reproduced). Brussels, 1937 Oil on canvas Museum Boijmans Van Beuningen, Rotterdam Narrator: Edward James, a British poet and patron of Surrealism, commissioned Magritte to make this portrait in 1937, titled Not to be Reproduced. Curator, Art Institute of Chicago, Stephanie D’Alessandro: When you look at James’s head you see that hair, and every lock is so carefully described—and the beautiful softness of his flesh and the hard highlight of the collar and the wrinkles in his suit. Narrator: James stands before a mirror—but Magritte does not reveal the reflection of his face. Stephanie D’Alessandro: It’s a very frustrating experience, in a way, and wonderful at the same time it’s part of his larger project about what painting traditionally is supposed to tell us, especially painting that is in a naturalistic mode. The whole idea of a title about reproduction is interesting here. We think about reproduction as something that tells us the truth. Reproduction also suggests multiplicity. All of that plays into the image that we see on this canvas. Narrator: On the mantel is Edgar Allan Poe’s novel, The Narratives of Arthur Gordon Pym. Stephanie D’Alessandro: Poe looms large for Magritte. Not only as someone who evokes a kind of disquieting mood, but also someone who plays with narrative in interesting ways, leads people down uncomfortable paths. — Learn more about this exhibition Top Privacy policyTerms of useUse high-contrast text © 2021 The Museum of Modern Art 0:01 1:15 17 / 18 be Reproduced” |
In art, Magritte’s painting “Not to be Reproduced” [Fig.6] depicting a man seeing himself in a mirror from behind, expresses the sense of the observer and the observed, while Henry Moore confronts us with the question as to whether the negative space in his sculptures has equal or greater value than the granite or bronze? With the merging of evolution and topology there is the opportunity to solve these enigmas scientifically and mathematically at long last.



 The Future of Humanity Is Our Responsibility
The Future of Humanity Is Our Responsibility 